两系法杂交稻是我国首创的水稻杂种优势利用技术,与三系法相比,两系法具有一系两用的特点,简化了育种流程。两用核不育系不受恢保关系限制,配组自由,稻种资源利用率高、选配优良组合机率高,成为中国水稻杂种优势利用的重要途径,极大推动了杂交水稻的发展。最新的农村农业部全国农作物推广数据显示两系杂交水稻种植面积占到杂交水稻总面积的50%左右,其中超过95%的温敏两系杂交稻组合都是由含有温敏雄性不育基因tms5的不育系配组而成,凸显了tms5在两系法杂交稻育种中的核心地位。
前期,中国科学院遗传与发育生物学研究所曹晓风团队和华南农业大学庄楚雄团队合作成功克隆了TMS5基因,该基因编码保守的核糖核酸酶RNase ZS1 (Zhou et al., 2014, Nature Communications)。近期,遗传发育所曹晓风团队、陈宇航团队和袁隆平农业高科技股份有限公司杨远柱团队合作揭示了TMS5作为tRNA环磷酸酶修复2′,3′-环磷-tRNA调节tRNA的循环进而调控水稻温敏雄性不育的分子机制 (Yan et al., 2024, Cell Research)。然而,tms5介导的温敏雄性不育的分子调控网络仍有待进一步阐明。近日,该团队揭示了E3泛素连接酶OsHel2通过阻碍停滞核糖体上mRNA的通读,来调节温敏雄性不育水稻的育性转换,相关成果于2024年11月13日以The E3 ligase OsHel2 impedes readthrough of stalled mRNAs to regulate male fertility in thermo-sensitive genic male sterile rice为题在Plant Communications上在线发表。
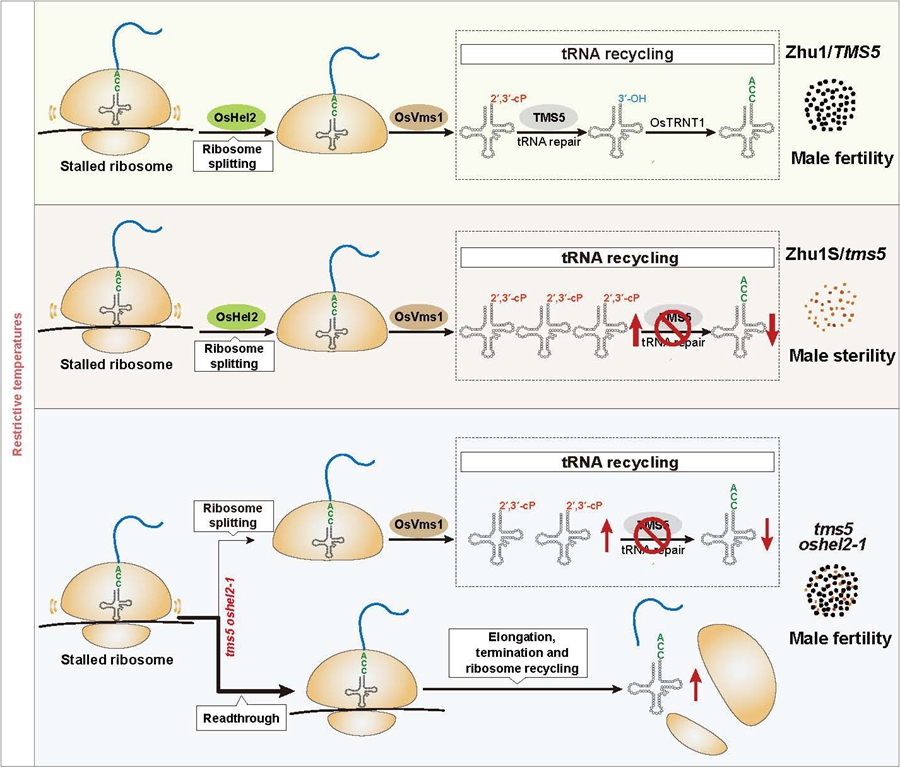
研究团队通过EMS诱变筛选获得一个中高温下可以恢复株1S(国内配制杂交早稻品种最多的温敏核不育系,携带tms5突变位点)育性抑制子sot3。遗传分析和高通量测序显示SOT3编码一个RING类型的E3泛素连接酶,抑制子中的SOT3蛋白在N端RING结构域的重要酶活位点发生了点突变(第29位Cys突变为Ser,C29S)。由于SOT3和酵母Hel2蛋白同源,因此我们将其命名为OsHel2。遗传互补和基因编辑实验证实OsHel2可以调控tms5突变体的育性转换过程。体外酶活和酶活点突变转基因实验结果表明,OsHel2的E3泛素连接酶活性对于其生物学功能至关重要。前期,研究团队分别开发了高通量测定环磷RNA(RcP-RNA-seq)和全长tRNA(FINE-tRNA-seq)的技术,研究发现tms5突变体株1S中由于失去环磷tRNA修复酶活,导致环磷tRNA过度积累,成熟tRNA丰度降低,揭示了成熟tRNA缺乏是导致tms5突变体高温不育的主要原因(Yan et al., 2024)。本研究发现抑制子tms5 oshel2-1中环磷tRNA和成熟tRNA的表达水平都较大程度地恢复到了野生型水平。为了进一步研究OsHel2的生物学功能,研究团队在水稻原生质体中建立了一种可在单细胞分辨率下定量检测细胞内核糖体停滞程度的双荧光报告系统,结果显示poly (A)序列可以引发严重的核糖体停滞现象,抑制子tms5 oshel2-1中OsHel2的RING结构域关键酶活位点的突变(C29S)可以部分抑制poly (A)引发的翻译停滞,表明OsHel2在核糖体质量控制(RQC)通路过程中可以阻碍停滞的核糖体上mRNA的通读,从而确保RQC顺利进行。在抑制子tms5 oshel2-1中,由于OsHel2蛋白发生突变,只有小部分停滞的核糖体进入RQC通路,环磷tRNA水平大幅降低。大多数停滞的核糖体可以进行正常的翻译和核糖体再循环过程。解离下来的tRNA弥补了成熟tRNA水平的不足,减轻了由于tms5突变造成的环磷tRNA不能修复的负担,使得抑制子育性得以恢复。
综上,该研究揭示了E3连接酶OsHel2通过阻碍停滞核糖体上mRNA的翻译,与TMS5协同调控温敏雄性不育水稻育性转换过程,建立了RQC成员的生物学功能与分子机制之间的直接联系。这些结果为与RQC成员相关的其他植物或动物的生理性疾病或缺陷的研究提供了新的思路。
曹晓风研究组已毕业客座博士研究生刘伟、已毕业博士研究生李吉、副研究员孙婧、副研究员刘春艳和助理研究员晏斌为该论文共同第一作者,曹晓风研究员、杨远柱研究员和刘春艳副研究员为共同通讯作者。宋显伟青年研究员参与了该项研究。该研究得到了农业农村部重大项目、国家重点研发计划和国家自然科学基金项目的资助。
图:OsHel2调控温敏不育水稻育性转换的分子机理模式图