韩方普研究组与合作者完成玉米B染色体精细图谱和功能研究
发布时间:2021.06.07
早在一个多世纪前发现B染色体在很多植物、动物和真菌基因组中广泛存在。B染色体对于个体的生命活动来说不是必需的,但它们仍然通过不同的机制存在于种群中。例如玉米B染色体不与任何A染色体配对,其传递不遵循孟德尔遗传定律,花粉第二次有丝分裂B染色体会发生不分离(nondisjunction),包含B染色体的精核优先与卵细胞受精,使其能够在群体中传递与累积。早期的玉米遗传学家利用A-B染色体易位系统创制丰富的染色体变异材料,广泛应用于玉米遗传学、人工染色体、基因剂量效应和着丝粒研究。例如在玉米中发现失活的B着丝粒可以稳定遗传(Han et al., PNAS, 2006);失活的B着丝粒仍然具有不分离的功能,并可以恢复活性(Han et al., Plant Cell, 2007;2009);利用B着丝粒与A染色体易位系探讨麦克林托克多年前提出的BFB理论(Birchler and Han, Plant Cell, 2018);利用B着丝粒错分裂材料详细研究新着丝粒形成(Zhang et al., Plant Cell, 2013;Liu et al., PNAS 2015)。B染色体为植物染色体生物学的发展做出了重要贡献,然而关于玉米B染色体的起源、进化及其在群体中积累的分子机制等关键问题仍然鲜为人知。
美国密苏里大学James A. Birchler研究组、捷克科学院实验植物研究所Jan Bartos研究组以及中国科学院遗传与发育生物学研究所韩方普研究组经过多年合作,共同完成玉米B染色体的基因组图谱,解析了玉米B染色体的起源、进化及不分离的机制。研究团队利用染色体分选、Illumina测序、Bionano光学图谱、Hi-C等方法相结合组装出328个B染色体特异的scaffolds,总长度125.9 Mb。随后利用B-A染色体易位、B着丝粒错分裂和B染色体断裂等特殊材料,构建染色体水平的分子图谱,注释出758个蛋白质编码基因,其中至少有88个基因表达。B染色体蛋白编码基因的同源物广泛散布在玉米10条A染色体上,但是A染色体没有检测到B染色体共线性的基因区域,推测当前B染色体的基因是长期进化过程由A染色体连续转移,随后发生部分降解的结果,这些保留下来的基因代表经过选择参与B染色体的功能维持,例如和“着丝粒区域”、“纺锤丝微管”、“胞质分裂的调节”、“组蛋白丝氨酸磷酸化”以及“染色体分离”等相关。进一步通过转座元件的分析发现玉米B染色体60%的序列由转座元件组成,其类型和A染色体基本相同。B染色体基因和转座元件的含量以及对转座蛋白编码基因的选择分析表明B染色体在进化世代中已经存在数百万年。
B染色体鉴定到574-kb的功能着丝粒序列,除了和A着丝粒共享CentC和CRM类型的重复序列外,还包含B着丝粒特异的串联重复序列B-repeat。着丝粒结构分析表明B着丝粒CENH3核小体的结构和功能与A着丝粒类似。高质量的B染色体序列图谱和遗传分析揭示B染色体不分离相关的顺式因子散布在着丝粒及其周围B染色体特异的重复序列上。通过B-3Sb缺失mapping,鉴定到B染色体末端34个蛋白编码基因可能作为反式作用元件参与不分离过程。B染色体转座重复基因可能进化出许多新功能,以维持种群中这种非必须染色体的存在。玉米B染色体精细图谱为进一步深入研究B染色体的特性提供了参考信息。
该论文于2021年6月5日在线发表于PNAS(DOI:10.1073/pnas.2104254118),Nicolas Blavet、杨华、苏汉东博士为该论文的共同第一作者,James A. Birchler、Jan Barto?和韩方普研究员为共同通讯作者。本工作得到国家自然科学基金委重点国际合作研究项目的支持。
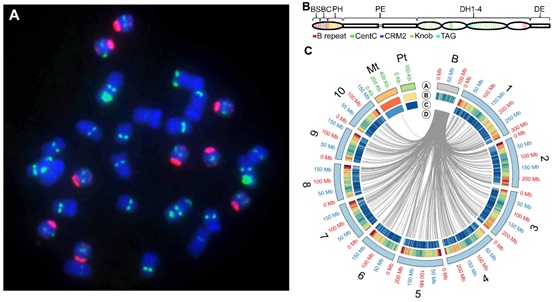
图: 玉米B染色体精细图谱和功能研究