小麦与黑麦的杂交工作始于19世纪70年代,英国A. S. Wilson以小麦为母本、黑麦为父本进行属间杂交获得真正的属间杂种,杂种高度不育。1888年,德国育种家W. Rimpau在普通小麦与黑麦的杂种不育株的一个穗子上得到种子,长成的植株能自行繁殖得到后代,这是由于低温使杂种F1自然加倍而形成的可育小黑麦。育种家和遗传学家看到小黑麦的优良性状,一百多年来,一直进行小麦与小黑麦的回交、自交来进行新品种选育。全世界现在推广的普通小麦品种中大约有70%含有一对特殊的染色体:我们称为1BL/1RS易位系,也就是普通小麦第一同源群B组染色体的长臂与黑麦的1R染色体短臂连接在一起形成小麦-黑麦染色体易位系。我国许多大面积推广的小麦新品种含有这条融合着丝粒的染色体。为什么这条易位染色体且是整臂易位在小麦育种中发挥了如此巨大的作用?
韩方普研究组长期从事小麦远缘杂交及染色体工程育种研究,从2009年开始,对全世界推广的几百份1B/1R易位系进行了染色体着丝粒结构研究,发现了非常有趣的结果:1B/1R易位系全部含有融合着丝粒,50%来自小麦,50%来自黑麦 (图A)。通过对黑麦着丝粒特异反转座子 (centromere specific retrotransposon) (pAWCR1) 的深入研究,发现黑麦的着丝粒反转座子在易位系小麦新品种中是活跃转录的 (图B)。麦类的CENH3基因位于第一同源群染色体的长臂 (Yuan et al, 2015, New Phytologist),CENH3组蛋白loading结果表明:易位系的黑麦部分着丝粒利用小麦的CENH3组蛋白行使着丝粒的功能。研究组也试图回答一个问题:为什么只有融合的着丝粒应用生产?多年努力想获得含有小麦着丝粒或黑麦着丝粒的1B/1R易位系,来研究转录及表达的影响,但几千份易位系筛选得到第一同源群易位系均是融合着丝粒易位染色体。
随着1RS抗性消失及育种需求,韩方普研究组利用从美国引进的200份黑麦材料进行新的小麦-黑麦杂交,获得大量双二倍体,并且完整建立了新的黑麦附加系和双端体系列材料,新的1B/1R易位系均是融合着丝粒。在1R的单体附加系后代,发现1R染色体片段或整臂可以易位到小麦1-7同源群,得到大量的染色体变异及序列变异新材料,包括着丝粒序列扩增及新着丝粒形成 (Guo et al, 2016, PLOS Genetics)。本工作为小麦育种提供了新的种质资源,也为染色体结构和功能研究提供了新的素材。根据ChIP-seq及基因组信息,初步进行易位系形成机制研究。新的易位系材料已经进行了回交转育工作,部分高代稳定品系已经进行不同区域观察及纳入育种系统工作 (图C)。
部分研究结果于2017年3月31日在Plant Journal在线发表。博士生王婧为第一作者。本项工作得到中科院战略先导专项和科技部资助。
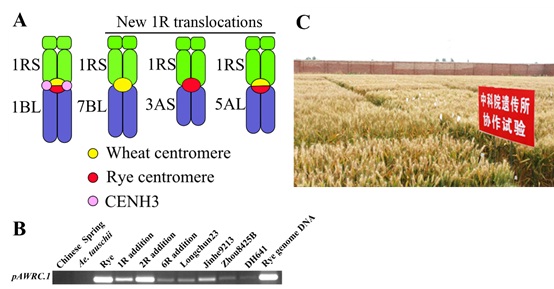
图. 1B/1R易位系着丝粒结构功能研究及回交转育。
(A): 1B/1R易位系着丝粒结构功能及具有不同着丝粒结构1R易位系染色体模式图;(B): RT-PCR检测1BL/1RS易位系中黑麦特异着丝粒序列pAWRC.1的转录情况;(C): 对1BL/1RS易位系进行回交转育及区域观察。
原文链接

